
35EXOc3'-5' exonuclease |
|---|
| SMART accession number: | SM00474 |
|---|---|
| Description: | 3\' -5' exonuclease proofreading domain present in DNA polymerase I, Werner syndrome helicase, RNase D and other enzymes |
| Interpro abstract (IPR002562): | This domain is responsible for the 3'-5' exonuclease proofreading activity of Escherichia coli DNA polymerase I (polI) and other enzymes, it catalyses the hydrolysis of unpaired or mismatched nucleotides. This domain consists of the amino-terminal half of the Klenow fragment in E. coli polI it is also found in the Werner syndrome helicase (WRN), focus forming activity 1 protein (FFA-1) and ribonuclease D (RNase D) [ (PUBMED:9697700) ]. Werner syndrome is a human genetic disorder causing premature aging; the WRN protein has helicase activity in the 3'-5' direction [ (PUBMED:9288107) (PUBMED:9224595) ]. The FFA-1 protein is required for formation of a replication foci and also has helicase activity; it is a homologue of the WRN protein [ (PUBMED:9697700) ]. RNase D is a 3'-5' exonuclease involved in tRNA processing. Also found in this family is the autoantigen PM/Scl thought to be involved in polymyositis-scleroderma overlap syndrome. |
| GO process: | nucleobase-containing compound metabolic process (GO:0006139) |
| GO function: | nucleic acid binding (GO:0003676), 3'-5' exonuclease activity (GO:0008408) |
| Family alignment: |
There are 40504 35EXOc domains in 40484 proteins in SMART's nrdb database.
Click on the following links for more information.
- Evolution (species in which this domain is found)
-
Taxonomic distribution of proteins containing 35EXOc domain.
This tree includes only several representative species. The complete taxonomic breakdown of all proteins with 35EXOc domain is also avaliable.
Click on the protein counts, or double click on taxonomic names to display all proteins containing 35EXOc domain in the selected taxonomic class.
- Cellular role (predicted cellular role)
-
Cellular role: replication
- Literature (relevant references for this domain)
-
Primary literature is listed below; Automatically-derived, secondary literature is also avaliable.
- Brautigam CA, Sun S, Piccirilli JA, Steitz TA
- Structures of normal single-stranded DNA and deoxyribo-3'-S-phosphorothiolates bound to the 3'-5' exonucleolytic active site of DNA polymerase I from Escherichia coli.
- Biochemistry. 1999; 38: 696-704
- Display abstract
The interaction of a divalent metal ion with a leaving 3' oxygen is a central component of several proposed mechanisms of phosphoryl transfer. In support of this are recent kinetic studies showing that thiophilic metal ions (e.g., Mn2+) stimulate the hydrolysis of compounds in which sulfur takes the place of the leaving oxygen. To examine the structural basis of this phenomenon, we have solved four crystal structures of single-stranded DNA's containing either oxygen or sulfur at a 3'-bridging position bound in conjunction with various metal ions at the 3'-5' exonucleolytic active site of the Klenow fragment (KF) of DNA polymerase I from Escherichia coli. Two structures of normal ssDNA bound to KF in the presence of Zn2+ and Mn2+ or Zn2+ alone were refined at 2.6- and 2.25-A resolution, respectively. They serve as standards for comparison with other Mn2+- and Zn2+-containing structures. In these cases, Mn2+ and Zn2+ bind at metal ion site B in a nearly identical position to Mg2+ (Brautigam and Steitz (1998) J. Mol. Biol. 277, 363-377). Two structures of KF bound to a deoxyoligonucleotide that contained a 3'-bridging sulfur at the scissile phosphate were refined at 2.03-A resolution. Although the bridging sulfur compounds bind in a manner very similar to that of the normal oligonucleotides, the presence of the sulfur changes the metal ion binding properties of the active site such that Mn2+ and Zn2+ are observed at metal ion site B, but Mg2+ is not. It therefore appears that the ability of the bridging sulfur compounds to exclude nonthiophilic metal ions from metal ion site B explains the low activity of KF exonuclease on these substrates in the presence of Mg2+ (Curley et al. (1997) J. Am. Chem. Soc. 119, 12691-12692) and that the 3'-bridging atom of the substrate is influencing the binding of metal ion B prior to catalysis.
- Moser MJ, Holley WR, Chatterjee A, Mian IS
- The proofreading domain of Escherichia coli DNA polymerase I and other DNA and/or RNA exonuclease domains.
- Nucleic Acids Res. 1997; 25: 5110-8
- Display abstract
Prior sequence analysis studies have suggested that bacterial ribonuclease (RNase) Ds comprise a complete domain that is found also in Homo sapiens polymyositis-scleroderma overlap syndrome 100 kDa autoantigen and Werner syndrome protein. This RNase D 3'-->5' exoribonuclease domain was predicted to have a structure and mechanism of action similar to the 3'-->5' exodeoxyibonuclease (proofreading) domain of DNA polymerases. Here, hidden Markov model (HMM) and phylogenetic studies have been used to identify and characterise other sequences that may possess this exonuclease domain. Results indicate that it is also present in the RNase T family; Borrelia burgdorferi P93 protein, an immunodominant antigen in Lyme disease; bacteriophage T4 dexA and Escherichia coli exonuclease I, processive 3'-->5' exodeoxyribonucleases that degrade single-stranded DNA; Bacillus subtilis dinG, a probable helicase involved in DNA repair and possibly replication, and peptide synthase 1; Saccharomyces cerevisiae Pab1p-dependent poly(A) nuclease PAN2 subunit, required for shortening mRNA poly(A) tails; Caenorhabditis elegans and Mus musculus CAF1, transcription factor CCR4-associated factor 1; Xenopus laevis XPMC2, prevention of mitotic catastrophe in fission yeast; Drosophila melanogaster egalitarian, oocyte specification and axis determination, and exuperantia, establishment of oocyte polarity; H.sapiens HEM45, expressed in tumour cell lines and uterus and regulated by oestrogen; and 31 open reading frames including one in Methanococcus jannaschii . Examination of a multiple sequence alignment and two three-dimensional structures of proofreading domains has allowed definition of the core sequence, structural and functional elements of this exonuclease domain.
- Braithwaite DK, Ito J
- Compilation, alignment, and phylogenetic relationships of DNA polymerases.
- Nucleic Acids Res. 1993; 21: 787-802
- Ollis DL, Brick P, Hamlin R, Xuong NG, Steitz TA
- Structure of large fragment of Escherichia coli DNA polymerase I complexed with dTMP.
- Nature. 1985; 313: 762-6
- Display abstract
The 3.3-A resolution crystal structure of the large proteolytic fragment of Escherichia coli DNA polymerase I complexed with deoxythymidine monophosphate consists of two domains, the smaller of which binds zinc-deoxythymidine monophosphate. The most striking feature of the larger domain is a deep crevice of the appropriate size and shape for binding double-stranded B-DNA. A flexible subdomain may allow the enzyme to surround completely the DNA substrate, thereby allowing processive nucleotide polymerization without enzyme dissociation.
- Metabolism (metabolic pathways involving proteins which contain this domain)
-
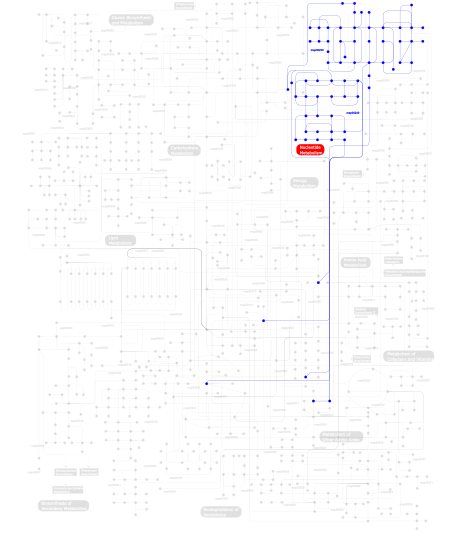
Click the image to view the interactive version of the map in iPath% proteins involved KEGG pathway ID Description 33.33  map00240
map00240Pyrimidine metabolism 33.33 map03030 DNA replication 33.33 map00230Purine metabolism This information is based on mapping of SMART genomic protein database to KEGG orthologous groups. Percentage points are related to the number of proteins with 35EXOc domain which could be assigned to a KEGG orthologous group, and not all proteins containing 35EXOc domain. Please note that proteins can be included in multiple pathways, ie. the numbers above will not always add up to 100%.
- Structure (3D structures containing this domain)
3D Structures of 35EXOc domains in PDB
PDB code Main view Title 1d8y 
CRYSTAL STRUCTURE OF THE COMPLEX OF DNA POLYMERASE I KLENOW FRAGMENT WITH DNA 1d9d 
CRYSTALL STRUCTURE OF THE COMPLEX OF DNA POLYMERASE I KLENOW FRAGMENT WITH SHORT DNA FRAGMENT CARRYING 2'-0-AMINOPROPYL-RNA MODIFICATIONS 5'-D(TCG)-AP(AUC)-3' 1d9f 
CRYSTAL STRUCTURE OF THE COMPLEX OF DNA POLYMERASE I KLENOW FRAGMENT WITH DNA TETRAMER CARRYING 2'-O-(3-AMINOPROPYL)-RNA MODIFICATION 5'-D(TT)-AP(U)-D(T)-3' 1dpi 
STRUCTURE OF LARGE FRAGMENT OF ESCHERICHIA COLI DNA POLYMERASE I COMPLEXED WITH D/TMP 1kfd 
CRYSTAL STRUCTURES OF THE KLENOW FRAGMENT OF DNA POLYMERASE I COMPLEXED WITH DEOXYNUCLEOSIDE TRIPHOSPHATE AND PYROPHOSPHATE 1kfs 
DNA POLYMERASE I KLENOW FRAGMENT (E.C.2.7.7.7) MUTANT/DNA COMPLEX 1kln 
DNA POLYMERASE I KLENOW FRAGMENT (E.C.2.7.7.7) MUTANT/DNA COMPLEX 1krp 
DNA POLYMERASE I KLENOW FRAGMENT (E.C.2.7.7.7) MUTANT/DNA COMPLEX 1ksp 
DNA POLYMERASE I KLENOW FRAGMENT (E.C.2.7.7.7) MUTANT/DNA COMPLEX 1l3s 
Crystal Structure of Bacillus DNA Polymerase I Fragment complexed to 9 base pairs of duplex DNA. 1l3t 
Crystal Structure of Bacillus DNA Polymerase I Fragment product complex with 10 base pairs of duplex DNA following addition of a single dTTP residue 1l3u 
Crystal Structure of Bacillus DNA Polymerase I Fragment product complex with 11 base pairs of duplex DNA following addition of a dTTP and a dATP residue. 1l3v 
Crystal Structure of Bacillus DNA Polymerase I Fragment product complex with 15 base pairs of duplex DNA following addition of dTTP, dATP, dCTP, and dGTP residues. 1l5u 
Crystal Structure of Bacillus DNA Polymerase I Fragment product complex with 12 base pairs of duplex DNA following addition of a dTTP, a dATP, and a dCTP residue. 1lv5 
Crystal Structure of the Closed Conformation of Bacillus DNA Polymerase I Fragment Bound to DNA and dCTP 1njw 
GUANINE-THYMINE MISMATCH AT THE POLYMERASE ACTIVE SITE 1njx 
THYMINE-GUANINE MISMATCH AT THE POLYMERASE ACTIVE SITE 1njy 
THYMINE-THYMINE MISMATCH AT THE POLYMERASE ACTIVE SITE 1njz 
CYTOSINE-THYMINE MISMATCH AT THE POLYMERASE ACTIVE SITE 1nk0 
ADENINE-GUANINE MISMATCH AT THE POLYMERASE ACTIVE SITE 1nk4 
GUANINE-GUANINE MISMATCH AT THE POLYMERASE ACTIVE SITE 1nk5 
ADENINE-ADENINE MISMATCH AT THE POLYMERASE ACTIVE SITE 1nk6 
CYTOSINE-CYTOSINE MISMATCH AT THE POLYMERASE ACTIVE SITE 1nk7 
GUANINE-ADENINE MISMATCH AT THE POLYMERASE ACTIVE SITE 1nk8 
A BACILLUS DNA POLYMERASE I PRODUCT COMPLEX BOUND TO A GUANINE-THYMINE MISMATCH AFTER A SINGLE ROUND OF PRIMER EXTENSION, FOLLOWING INCORPORATION OF DCTP. 1nk9 
A BACILLUS DNA POLYMERASE I PRODUCT COMPLEX BOUND TO A GUANINE-THYMINE MISMATCH AFTER TWO ROUNDS OF PRIMER EXTENSION, FOLLOWING INCORPORATION OF DCTP AND DGTP. 1nkb 
A BACILLUS DNA POLYMERASE I PRODUCT COMPLEX BOUND TO A GUANINE-THYMINE MISMATCH AFTER THREE ROUNDS OF PRIMER EXTENSION, FOLLOWING INCORPORATION OF DCTP, DGTP, AND DTTP. 1nkc 
A BACILLUS DNA POLYMERASE I PRODUCT COMPLEX BOUND TO A GUANINE-THYMINE MISMATCH AFTER FIVE ROUNDS OF PRIMER EXTENSION, FOLLOWING INCORPORATION OF DCTP, DGTP, DTTP, AND DATP. 1nke 
A BACILLUS DNA POLYMERASE I PRODUCT COMPLEX BOUND TO A CYTOSINE-THYMINE MISMATCH AFTER A SINGLE ROUND OF PRIMER EXTENSION, FOLLOWING INCORPORATION OF DCTP. 1qsl 
KLENOW FRAGMENT COMPLEXED WITH SINGLE-STRANDED SUBSTRATE AND EUROPIUM (III) ION 1u45 
8oxoguanine at the pre-insertion site of the polymerase active site 1u47 
cytosine-8-Oxoguanine base pair at the polymerase active site 1u48 
Extension of a cytosine-8-oxoguanine base pair 1u49 
Adenine-8oxoguanine mismatch at the polymerase active site 1u4b 
Extension of an adenine-8oxoguanine mismatch 1ua0 
Aminofluorene DNA adduct at the pre-insertion site of a DNA polymerase 1ua1 
Structure of aminofluorene adduct paired opposite cytosine at the polymerase active site. 1xc9 
Structure of a high-fidelity polymerase bound to a benzo[a]pyrene adduct that blocks replication 1xwl 
BACILLUS STEAROTHERMOPHILUS (NEWLY IDENTIFIED STRAIN AS YET UNNAMED) DNA POLYMERASE FRAGMENT 1yt3 
Crystal Structure of Escherichia coli RNase D, an exoribonuclease involved in structured RNA processing 2bdp 
CRYSTAL STRUCTURE OF BACILLUS DNA POLYMERASE I FRAGMENT COMPLEXED TO 9 BASE PAIRS OF DUPLEX DNA 2e6l 
structure of mouse WRN exonuclease domain 2e6m 
structure of mouse werner exonuclease domain 2fbt 
WRN exonuclease 2fbv 
WRN exonuclease, Mn complex 2fbx 
WRN exonuclease, Mg complex 2fby 
WRN exonuclease, Eu complex 2fc0 
WRN exonuclease, Mn dGMP complex 2hbj 
Structure of the yeast nuclear exosome component, Rrp6p, reveals an interplay between the active site and the HRDC domain 2hbk 
Structure of the yeast nuclear exosome component, Rrp6p, reveals an interplay between the active site and the HRDC domain; Protein in complex with Mn 2hbl 
Structure of the yeast nuclear exosome component, Rrp6p, reveals an interplay between the active site and the HRDC domain; Protein in complex with Mn, Zn, and AMP 2hbm 
Structure of the yeast nuclear exosome component, Rrp6p, reveals an interplay between the active site and the HRDC domain; Protein in complex with Mn, Zn, and UMP 2hhq 
O6-methyl-guanine:T pair in the polymerase-10 basepair position 2hhs 
O6-methyl:C pair in the polymerase-10 basepair position 2hht 
C:O6-methyl-guanine pair in the polymerase-2 basepair position 2hhu 
C:O6-methyl-guanine in the polymerase postinsertion site (-1 basepair position) 2hhv 
T:O6-methyl-guanine in the polymerase-2 basepair position 2hhw 
ddTTP:O6-methyl-guanine pair in the polymerase active site, in the closed conformation 2hhx 
O6-methyl-guanine in the polymerase template preinsertion site 2hvh 
ddCTP:O6MeG pair in the polymerase active site (0 position) 2hvi 
ddCTP:G pair in the polymerase active site (0 position) 2hw3 
T:O6-methyl-guanine pair in the polymerase postinsertion site (-1 basepair position) 2kfn 
KLENOW FRAGMENT WITH BRIDGING-SULFUR SUBSTRATE AND MANGANESE 2kfz 
KLENOW FRAGMENT WITH BRIDGING-SULFUR SUBSTRATE AND ZINC ONLY 2kzm 
KLENOW FRAGMENT WITH NORMAL SUBSTRATE AND ZINC AND MANGANESE 2kzz 
KLENOW FRAGMENT WITH NORMAL SUBSTRATE AND ZINC ONLY 2xo7 
Crystal structure of a dA:O-allylhydroxylamine-dC basepair in complex with fragment DNA polymerase I from Bacillus stearothermophilus 2xy5 
Crystal structure of an artificial salen-copper basepair in complex with fragment DNA polymerase I from Bacillus stearothermophilus 2xy6 
Crystal structure of a salicylic aldehyde basepair in complex with fragment DNA polymerase I from Bacillus stearothermophilus 2xy7 
Crystal structure of a salicylic aldehyde base in the pre-insertion site of fragment DNA polymerase I from Bacillus stearothermophilus 2y1i 
Crystal structure of a S-diastereomer analogue of the spore photoproduct in complex with fragment DNA polymerase I from Bacillus stearothermophilus 2y1j 
Crystal structure of a R-diastereomer analogue of the spore photoproduct in complex with fragment DNA polymerase I from Bacillus stearothermophilus 3bdp 
DNA POLYMERASE I/DNA COMPLEX 3cym 
Crystal structure of protein BAD_0989 from Bifidobacterium adolescentis 3eyz 
Cocrystal structure of Bacillus fragment DNA polymerase I with duplex DNA (open form) 3ez5 
Cocrystal structure of Bacillus fragment DNA polymerase I with duplex DNA , dCTP, and zinc (closed form). 3hp6 
Crystal structure of fragment DNA polymerase I from Bacillus stearothermophilus F710Y mutant bound to G:T mismatch 3hpo 
Crystal structure of fragment DNA polymerase I from Bacillus stearothermophilus Y714S mutant bound to G:T mismatch 3ht3 
Crystal structure of fragment DNA polymerase I from Bacillus stearothermophilus V713P mutant bound to G:dCTP 3pv8 
Crystal Structure of Bacillus DNA Polymerase I Large Fragment Bound to DNA and ddTTP-dA in Closed Conformation 3px0 
Crystal Structure of Bacillus DNA Polymerase I Large Fragment Bound to DNA and dCTP-dA Mismatch (tautomer) in Closed Conformation 3px4 
Crystal Structure of Bacillus DNA Polymerase I Large Fragment Bound to DNA and ddCTP-dA Mismatch (wobble) in Ajar Conformation 3px6 
Crystal Structure of Bacillus DNA Polymerase I Large Fragment Bound to DNA and ddCTP-dA Mismatch (tautomer) in Closed Conformation 3saf 
Crystal structure of the human RRP6 catalytic domain with D313N mutation in the active site 3sag 
Crystal structure of the human RRP6 catalytic domain with D313N mutation in the active site 3sah 
Crystal structure of the human RRP6 catalytic domain with Y436A mutation in the catalytic site 3tan 
Crystal Structure of Bacillus DNA Polymerase I Large Fragment Bound to Duplex DNA with Cytosine-Adenine Mismatch at (n-1) Position 3tap 
Crystal Structure of Bacillus DNA Polymerase I Large Fragment Bound to Duplex DNA with Cytosine-Adenine Mismatch at (n-3) Position 3taq 
Crystal Structure of Bacillus DNA Polymerase I Large Fragment Bound to Duplex DNA with Cytosine-Adenine Mismatch at (n-4) Position 3tar 
Crystal Structure of Bacillus DNA Polymerase I Large Fragment Bound to Duplex DNA with Cytosine-Adenine Mismatch at (n-6) Position 3thv 
Crystal Structure of Bacillus DNA Polymerase I Large Fragment Bound to DNA and ddATP-dT in Closed Conformation 3ti0 
Crystal Structure of Bacillus DNA Polymerase I Large Fragment Bound to DNA and ddGTP-dC in Closed Conformation 4b9l 
Structure of the high fidelity DNA polymerase I with the oxidative formamidopyrimidine-dA DNA lesion in the pre-insertion site. 4b9m 
Structure of the high fidelity DNA polymerase I with an oxidative formamidopyrimidine-dA DNA lesion -thymine basepair in the post- insertion site. 4b9n 
Structure of the high fidelity DNA polymerase I correctly bypassing the oxidative formamidopyrimidine-dA DNA lesion. 4b9s 
Structure of the high fidelity DNA polymerase I with an oxidative formamidopyrimidine-dG DNA lesion outside of the pre-insertion site. 4b9t 
Structure of the high fidelity DNA polymerase I with an oxidative formamidopyrimidine-dG DNA lesion -dC basepair in the post-insertion site. 4b9u 
Structure of the high fidelity DNA polymerase I with an oxidative formamidopyrimidine-dG DNA lesion -dA basepair in the post-insertion site. 4b9v 
Structure of the high fidelity DNA polymerase I with extending from an oxidative formamidopyrimidine-dG DNA lesion -dA basepair. 4bdp 
CRYSTAL STRUCTURE OF BACILLUS DNA POLYMERASE I FRAGMENT COMPLEXED TO 11 BASE PAIRS OF DUPLEX DNA AFTER ADDITION OF TWO DATP RESIDUES 4dqi 
Ternary complex of Bacillus DNA Polymerase I Large Fragment, DNA duplex, and dCTP (paired with dG of template) 4dqp 
Ternary complex of Bacillus DNA Polymerase I Large Fragment, DNA duplex, and ddCTP (paired with dG of template) 4dqq 
Ternary complex of Bacillus DNA Polymerase I Large Fragment E658A, DNA duplex, and rCTP (paired with dG of template) in presence of Mg2+ 4dqr 
Ternary complex of Bacillus DNA Polymerase I Large Fragment E658A, DNA duplex, and rCTP (paired with dG of template) in presence of Mn2+ 4dqs 
Binary complex of Bacillus DNA Polymerase I Large Fragment and duplex DNA with rC in primer terminus paired with dG of template 4ds4 
Ternary complex of Bacillus DNA Polymerase I Large Fragment, DNA duplex, and rCTP in presence of Mn2+ 4ds5 
Ternary complex of Bacillus DNA Polymerase I Large Fragment, DNA duplex, and rCTP in presence of Mg2+ 4dse 
Ternary complex of Bacillus DNA Polymerase I Large Fragment F710Y, DNA duplex, and rCTP (paired with dG of template) in presence of Mg2+ 4dsf 
Ternary complex of Bacillus DNA Polymerase I Large Fragment F710Y, DNA duplex, and rCTP (paired with dG of template) in presence of Mn2+ 4dsi 
Crystal structure of fragment DNA polymerase I from Bacillus stearothermophilus with duplex DNA, Se-dGTP and Calcium 4dsj 
Crystal structure of fragment DNA polymerase I from Bacillus stearothermophilus with duplex DNA, dGTP and Calcium 4dsk 
Crystal structure of fragment DNA polymerase I from Bacillus stearothermophilus with duplex DNA, PPi and Calcium 4dsl 
Crystal structure of fragment DNA polymerase I from Bacillus stearothermophilus with duplex DNA and Calcium 4dwi 
Crystal structure of fragment DNA polymerase I from Bacillus stearothermophilus with self complementary DNA, Se-dGTP and Calcium 4e0d 
Binary complex of Bacillus DNA Polymerase I Large Fragment E658A and duplex DNA 4ez6 
Bacillus DNA Polymerase I Large Fragment Complex 1 4ez9 
Bacillus DNA Polymerase I Large Fragment Complex 2 4f2r 
DNA Polymerase I Large Fragment complex 3 4f2s 
DNA Polymerase I Large Fragment complex 4 4f3o 
DNA Polymerase I Large Fragment Complex 5 4f4k 
DNA Polymerase I Large Fragment Complex 6 4f8r 
Bacillus DNA Polymerase I Large Fragment complex 7 4nlb 
Crystal structure of the catalytic core of RRP6 from Trypanosoma brucei 4nlc 
Crystal structure of the catalytic core of RRP6 from Trypanosoma brucei, mutant C496S 4o0i 
4O0I 4oo1 
4OO1 4uqg 
4UQG 4yfu 
4YFU 5c0w 
5C0W 5c0x 
5C0X 5c0y 
5C0Y 5dkt 
5DKT 5dku 
5DKU 5fiq 
5FIQ 5fis 
5FIS 5k36 
5K36 - Links (links to other resources describing this domain)
-
PFAM 3_5_exonuclease INTERPRO IPR002562